Wat betreft de waterflora voldoet de ecologische kwaliteit van veel KRW-waterlichamen nog niet. De vraag is welke aangrijpingspunten waterschappen hebben om deze kwaliteit te verbeteren. Door vegetatieopnames te koppelen aan fysisch-chemische en hydromorfologische data is een aantal duidelijke sleutelfactoren geïdentificeerd. Verrassend is dat ammonium een sterkere relatie blijkt te hebben met de soortensamenstelling dan nitraat. Daarnaast is belangrijk dat herstel van stroming en afvoer in beken aanzienlijke invloed heeft op de waterflora.
Download hier de pdf van dit artikel.
Op basis van de waterflora kreeg in 2014 37% van de KRW-wateren de beoordeling ‘onvoldoende’ of ‘slecht’ [1]. Op veel plaatsen zijn dus maatregelen nodig om deze kwaliteit te verbeteren. Maatregelen zijn pas effectief als ze aangrijpen op milieufactoren die de soortensamenstelling beïnvloeden. Om inzicht te krijgen in deze sleutelfactoren hebben de waterschappen Rijn en IJssel en Vallei en Veluwe in 2014 hun gegevens van waterflora en abiotiek nader geanalyseerd [2].
Gebruikte gegevens
Voor de analyses hebben we gebruik gemaakt van vegetatieopnames: 70 in laaglandbeken (KRW-typen R4 en R5) en 47 in lijnvormige watergangen met meestal stilstaand water (KRW-typen M3 en M6). De opnames, gemaakt volgens de KRW-systematiek in de periode 2007-2013, zijn gerelateerd aan fysisch-chemische factoren van de waterlaag en aan afmetingen en afvoerregime van de watergangen. Als van een locatie meerdere opnames uit deze periode beschikbaar zijn, dan is de meest recente gebruikt. Aangezien licht cruciaal is voor waterplanten, zijn de beboste locaties met meer dan 15% beschaduwing buiten beschouwing gelaten. Fysisch-chemische gegevens van een locatie zijn verzameld in hetzelfde jaar als de vegetatieopname of in het eraan voorafgaande jaar. Voor de alkaliniteit moesten we soms terugvallen op oudere data omdat dit geen standaardonderdeel is van de monitoring. Wat betreft het afvoerregime is voor iedere locatie vastgesteld of er het gehele jaar afvoer is of dat er periodiek sprake is van afvoerloosheid of droogval. De stroomsnelheden zijn geschat op basis van de omvang van de jaarlijkse piekafvoeren en de dimensie van de watergang. Hoewel het maaibeheer van de watergangen ook belangrijk is [3], hebben we deze factor niet in de analyse meegenomen aangezien er amper verschillen waren tussen de watergangen in tijdstip en frequentie van maaien.
Sleutelfactoren op het spoor
In de opnamen kwamen 145 soorten macrofyten voor, waarvan liesgras (in 85% van de opnamen), klein kroos (80%) en smalle waterpest (78%) het meest frequent aanwezig waren. Met het programma TWINSPAN [4] zijn de opnamen ingedeeld in tien soortenclusters verdeeld over twee hoofdgroepen: 1) met gele plomp en pijlkruid, voorkomend in brede, diepe wateren en 2) met mannagras, voorkomend in smalle, ondiepe wateren (afbeelding 1).
De relaties tussen soorten en milieufactoren zijn geanalyseerd met een canonieke correspondentie-analyse (CCA) [5]. Met deze techniek zijn tegelijkertijd de relaties tussen meerdere soorten en meerdere abiotische variabelen te onderzoeken. De breedte van de watergang, in combinatie met stroomsnelheid en afvoerloosheid, alkaliniteit, fosfor en stikstof verklaarde in totaal 32% van de variantie in soortensamenstelling. Deze factoren kwamen ook als belangrijkste uit de bus bij een studie naar waterplanten in Deense laaglandbeken [3]. Afbeelding 1 presenteert de kenmerkende soorten van de clusters en hun eigenschappen in termen van twee sleutelfactoren: stikstof en alkaliniteit.
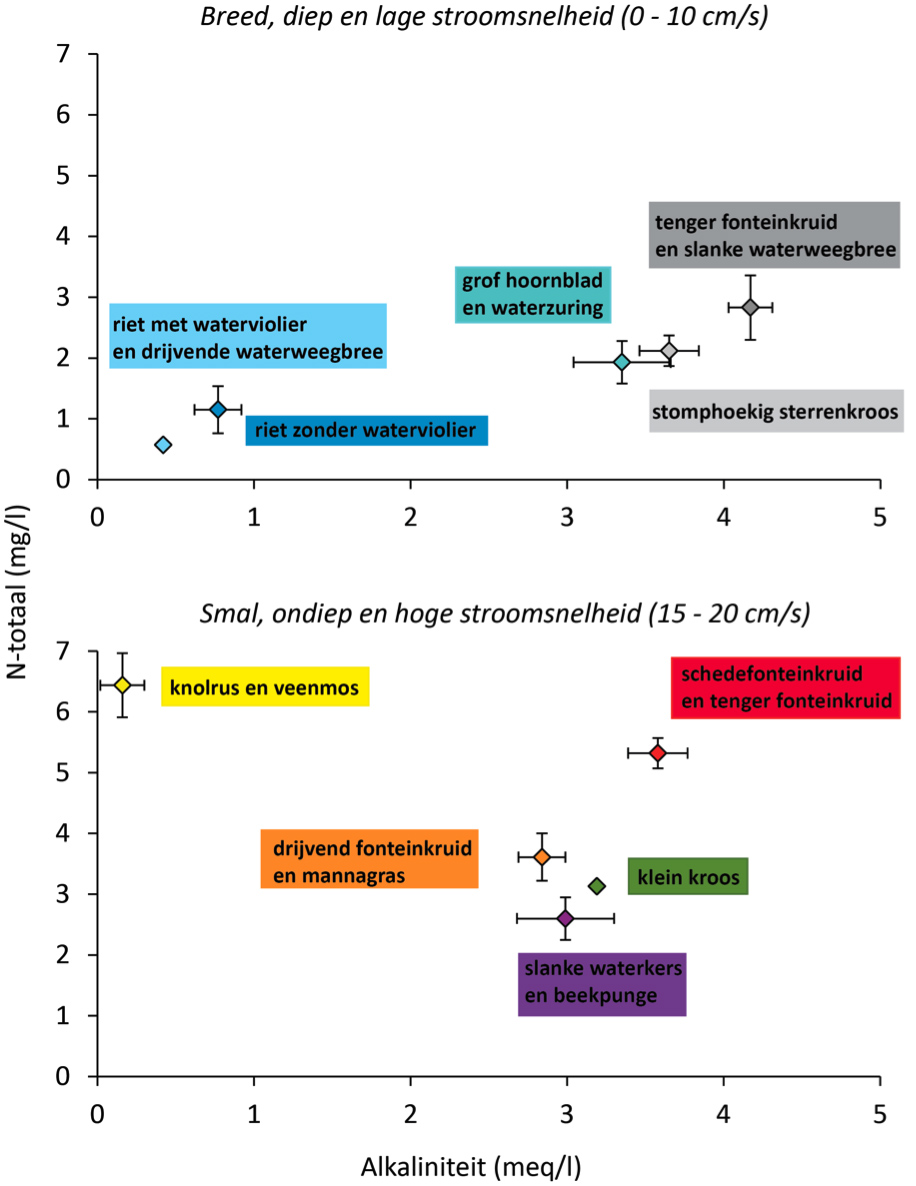
Afbeelding 1. Positie van de TWINSPAN-clusters van water- en oeverplanten (aangeduid met hun kenmerkende soorten) op de gradiënten van alkaliniteit en totaal-stikstof
Per cluster is een gemiddelde (± standaardfout) van elke milieuvariabele berekend op basis van de waarde van iedere opname binnen zo’n cluster. De clusters waren duidelijk te onderscheiden in een groep met brede (bovenste paneel) en smalle watergangen (onderste paneel). Deze groepen verschilden ook in stroomsnelheid bij jaarlijkse piekafvoer (T=1).
Breedte en stroomsnelheid
Breedte is waarschijnlijk niet rechtstreeks bepalend voor water- en oeverplanten. Brede watergangen zijn echter meestal ook dieper, wat de mogelijkheden voor uitbreiding van oeverplanten en de lichtbeschikbaarheid voor waterplanten beperkt. Daarnaast was in ons onderzoek de stroomsnelheid meestal lager in brede watergangen. Hierdoor bezinkt meer slib, wat een belangrijk knelpunt voor soortenrijke vegetaties vormt [6].
Alkaliniteit
Alkaliniteit lijkt vooral bepalend voor het voorkomen van de groep met waterviolier en drijvende waterweegbree en de groep met knolrus en veenmos enerzijds (lage alkaliniteit) en de overige groepen anderzijds (hogere alkaliniteit) (afbeelding 1). De eerste twee groepen waren beperkt tot respectievelijk het Apeldoorns Kanaal en de Heelsumse beek, beide aan de rand van de Veluwe. Alkaliniteit bleef ook overeind als sleutelfactor in een afzonderlijke analyse van beken in de Achterhoek en Liemers. Kenmerkend voor wateren met lage alkaliniteit waren hier drijvend fonteinkruid en buigzaam kranswier. Stijve en fijne waterranonkel waren kenmerkend voor wateren met een hoge alkaliniteit. Wateren met een hoge alkaliniteit bevatten veel bicarbonaat, dat de afbraak van organisch materiaal in de bodem en daarmee de nutriëntenbeschikbaarheid stimuleert [7].
Nutriënten
Fosfor in de waterlaag kwam als sleutelfactor uit de bus in een CCA met jaargemiddelde, maar niet met zomergemiddelde concentraties. Een verklaring is dat in voedselrijke wateren in de zomer veel fosfaat in biomassa is opgenomen zodat wateren met uiteenlopende begroeiing een ogenschijnlijk overeenkomstige fosfaatconcentratie hebben [6]. De hoogste jaargemiddelde concentratie (0,36 mg/l P) werd gemeten in het cluster met schedefonteinkruid en grof hoornblad, de laagste (0,01 mg/l P) in de vegetatie met waterviolier en drijvende waterweegbree. Stikstof kwam zowel met jaar- als zomergemiddelde concentraties naar voren als sleutelfactor (zie hierna).
De ecologische kwaliteit in relatie tot milieufactoren
De ecologische kwaliteitsratio
De ecologische kwaliteitsratio (EKR) is de verhouding van de waargenomen waarde van een KRW-kwaliteitselement en de referentiewaarde van dat element, en hangt af van de soortensamenstelling en de hoeveelheid waterplanten in een watergang [8, 9]. Voor stromende wateren vonden we met behulp van meervoudige regressieanalyse geen relatie tussen nutriënten en EKR-score, maar er was wel een effect van afvoerloosheid (tabel 1).
Tabel 1. Resultaten van de meervoudige regressieanalyse voor de KRW-maatlatscore (EKR= Ecologische Kwaliteits Ratio) van beken en lijnvormige, gegraven watergangen en de abundantie van draadwier en kroos in beken
Voor de milieufactoren die een significant deel van de variatie verklaren zijn de gestandaardiseerde regressiecoëfficiënten weergegeven. Dit maakt het mogelijk om het relatieve effect ten opzichte van de andere significante factoren te beoordelen. R2 geeft weer welk deel van de variatie verklaard wordt door de (combinatie van) significante factor(en). Analyses van EKR zijn uitgevoerd na logit-transformatie van EKR [17] en log-transformatie van concentraties. Afvoerloosheid is een variabele met twee waarden: 1 voor stroming en 2 voor stilstand. Voor concentraties van stoffen zijn zomergemiddelden gebruikt.
De gemiddelde EKR (± standaardfout) van permanent stromende beken (0,36 ± 0,02; n = 37) is significant (P = 0,044) hoger dan die van beken met tijdelijke afvoerloosheid of droogval (0,31 ± 0,02; n = 33). De EKR van de andere watergangen is ook significant (P = 0,022) hoger in watergangen die ’s zomers doorstroming houden (0,44 ± 0,04; n = 33) dan in wateren die ’s zomers stilvallen (0,33 ± 0,02; n = 14). Daarnaast blijkt ammonium ook belangrijk (tabel 1): de EKR is lager bij hogere ammoniumconcentraties (afbeelding 2).
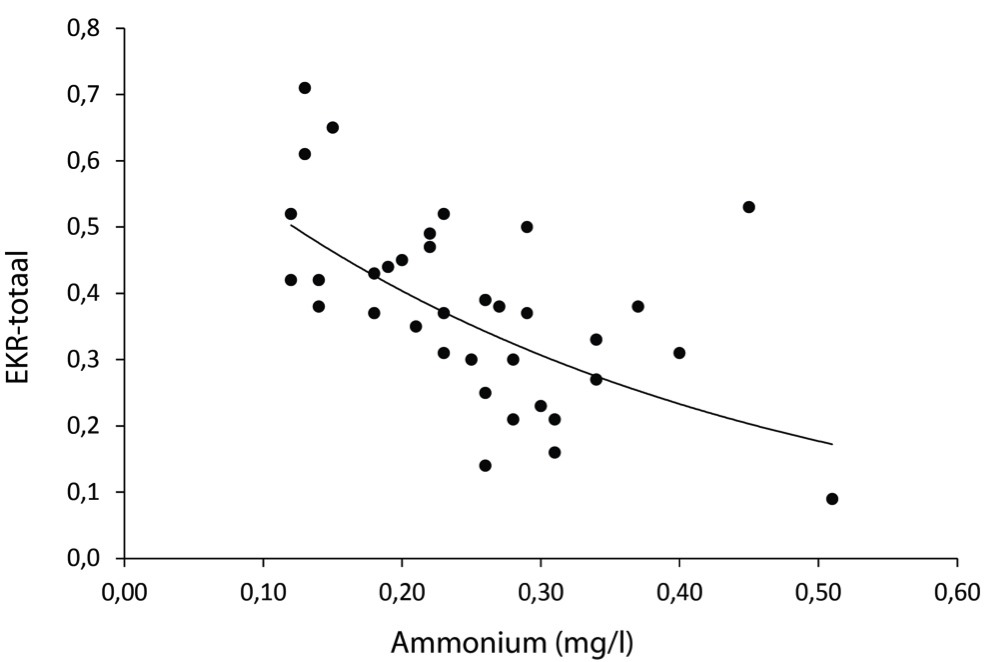
Afbeelding 2. Relatie tussen EKR-totaal en de zomergemiddelde ammoniumconcentratie voor lijnvormige, meestal stilstaande watergangen (typen M3 en M6)
In deze grafiek zijn vier extreme waarden (tussen 1 en 4 mg/l) weggelaten. Voor significantie zie tabel 1.
Die lage EKR is een gevolg van de hoge abundantie van schedefonteinkruid en grof hoornblad bij hoge ammoniumconcentraties (afbeelding 4). Mogelijk prefereren deze soorten ammonium boven nitraat als stikstofbron [10], waardoor ze bij hoge ammoniumconcentraties minder concurrentiekrachtige soorten verdringen. Daarnaast is ook een meer indirecte relatie met ammonium denkbaar. Ammonium wordt, met behulp van zuurstof, snel omgevormd naar nitraat. Veel ammonium kan dus leiden tot een lage zuurstofconcentratie. In plaats van oorzaak kan veel ammonium echter ook een gevolg zijn van lage zuurstofconcentraties; omvorming naar nitraat is dan immers niet meer mogelijk. In ieder geval duidt een hoge ammoniumconcentratie op gereduceerde omstandigheden en juist dan komt veel fosfaat uit de waterbodem vrij door sulfaatreductie [6]. Dat extra fosfaat zal vervolgens concurrentiekrachtige soorten als schedefonteinkruid en grof hoornblad bevoordelen.
Kroos en draadwieren
Lagen van kroos en draadwieren onderscheppen licht voor onderwaterplanten en zorgen bovendien voor een sterke zuurstofdaling ’s nachts [11]. In de KRW-beoordeling van de waterflora in beken nemen ze daarom een prominente plek in [9]. In beken blijkt de bedekking van klein kroos positief gecorreleerd te zijn met fosfaat, die van draadwier positief met sulfaat en ammonium, maar negatief met nitraat (tabel 1). De laatste relatie is het beste te verklaren door processen op het grensvlak van waterlaag en waterbodem, waar draadwier zich vooral ontwikkelt. Onder anaërobe condities wordt sulfaat door bacteriën in de waterbodem gereduceerd. Het sulfide dat hierbij ontstaat bindt het ijzer uit ijzerfosfaten in de bodem. Het gemobiliseerde fosfaat diffundeert vervolgens naar de waterlaag en komt als eerste beschikbaar voor draadwier op de bodem [6, 7]. Hoge concentraties nitraat voorkomen dat sulfaat gereduceerd wordt omdat bacteriën eerst alle beschikbare nitraat benutten [12]. In eerder onderzoek [11] bleken kroosdekken ook gestuurd te worden door fosfaat uit de waterbodem. Daarnaast is ook het afvoerregime bepalend voor kroos (tabel 1). Permanent stromende beken met hoge fosfaatconcentraties hebben een kortere verblijftijd en daardoor een minder sterke ontwikkeling van kroos dan zomers stilvallende beken.
Implicaties voor beheer
Drempelwaarden voor fosfor
Een zomergemiddelde totaal-P-concentratie van 0,12 mg/l lijkt een duidelijke grens voor de omslag van positief naar negatief gewaardeerde soorten (afbeelding 3). Deze concentratie komt overeen met de voorgestelde norm voor beken [9]. De norm voor niet-stromende, lijnvormige wateren (M3 en M6) is minder streng (0,15 mg/l) [13], maar onze resultaten geven aanleiding om die norm te heroverwegen.

Afbeelding 3. Gewogen zomergemiddelde concentraties van P en N waarbij waterplanten zijn aangetroffen
Gemiddelden zijn gewogen naar hun abundantie. Met rood zijn de soorten aangegeven die in de KRW-maatlatten voor R5 en M3 negatief en met blauw de soorten die positief scoren. Soorten die in de ene maatlat negatief en in de andere positief scoren worden voorgesteld door rode stippen met een blauwe rand. Open stippen zijn soorten zonder KRW-indicatiewaarde. Alleen soorten met 4 of meer waarnemingen zijn opgenomen. Soorten zijn aangeduid als afkortingen van hun wetenschappelijke naam: eerste drie letters van geslachtsnaam en eerste drie letters van soortnaam. Bijvoorbeeld: Pot.pec. = Potamogeton pectinatus (schedefonteinkruid).
Drempelwaarden voor stikstof
De meeste in de KRW-maatlat positief gewaardeerde soorten komen voor bij een zomergemiddelde N-totaal concentratie ≤ 2,7 mg/l (afbeelding 3), wat niet ver verwijderd is van de grens tussen ‘goed’ en ‘matig’ in beken (2,3 mg/l) [9].
Voor ammonium zijn normen geformuleerd voor het jaargemiddelde (0,304 mg/l bij pH 7,7 en T=15 oC) en de 90 percentiel (0,608 mg/l bij pH 7,7 en T=15 oC), waarden die in hooguit 10% van de jaarlijkse metingen overschreden mogen worden. Deze normen zijn gebaseerd op ecotoxicologisch onderzoek aan dieren [14]. Ammonium kan echter ook toxisch zijn voor onderwaterplanten [15], wat overigens afhangt van temperatuur, zuurgraad en lichtbeschikbaarheid. Zoals we hierboven hebben beargumenteerd, werkt ammonium verder als voedingsstof en heeft het invloed op de beschikbaarheid van andere voedingsstoffen. Het is daarom zinvol om de normen voor ammonium ook hierop en op zijn mogelijke toxiciteit voor onderwaterplanten af te stemmen. Op basis van onze data lijkt een zomergemiddelde concentratie van 0,2 mg NH4-N/l een passende norm omdat bij deze concentratie de scheidslijn ligt tussen positief en negatief gewaardeerde soorten; voor het jaargemiddelde is dit 0,3 mg/l (afbeelding 4).
Wat betreft de nitraatconcentratie is er geen verschil in voorkomen tussen soorten die een positieve of negatieve score op de KRW-maatlat geven: de meeste soorten komen voor bij een jaargemiddelde nitraatconcentratie ≤ 3,0 mg/l en een zomergemiddelde concentratie ≤ 2,2 mg/l (afbeelding 4).
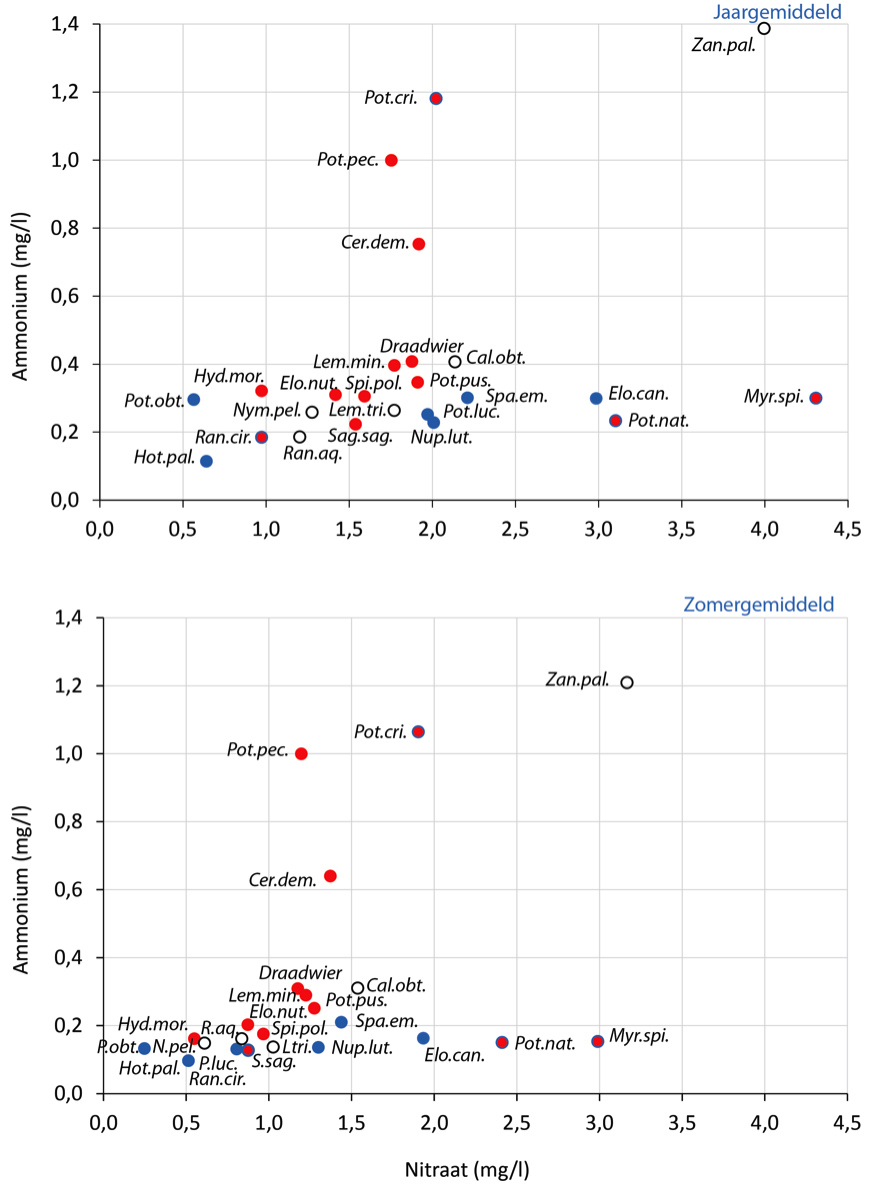
Figuur 4. Gewogen jaar- en zomergemiddelde concentraties van ammonium en nitraat waarbij waterplanten zijn aangetroffen
Voor toelichting en legenda, zie afbeelding 3.
Sturen op nitraat- en/of ammoniumreductie?
Waterschappen hebben zelf een belangrijke stuurknop voor ammonium in handen. In wateren die effluent ontvangen, is vaak een aanzienlijk deel van het ammonium afkomstig van rioolwaterzuiveringen. Door op de rioolwaterzuivering langer te beluchten kan de ammoniumuitstoot omlaag. Ammonium wordt dan omgezet in nitraat, dat vervolgens in het oppervlaktewater terechtkomt. Meer beluchten betekent dus ook meer nitraat in het oppervlaktewater accepteren. De vraag is of dit erger is dan een hogere ammoniumbelasting.
In onze studie lijkt nitraat een belemmering voor draadwier en is daarmee positief voor andere soorten. Dit positieve effect is waarschijnlijk een gevolg van remmende werking op sulfaatreductie. Een eventueel positief effect van nitraat is dus sterk context-afhankelijk: vooral bij hoge sulfaatconcentraties en organische waterbodems zijn positieve effecten van nitraat te verwachten. Er is echter ook experimenteel bewijs dat hoge nitraatconcentraties negatief kunnen uitpakken voor afzonderlijke soorten [16]. De rol van nitraat verdient meer onderzoek om een goede afweging te kunnen maken. Vooralsnog heeft het de voorkeur om in te zetten op lage ammoniumbelasting wanneer het niet mogelijk is de totale stikstofvracht te reduceren.
Implicaties voor monitoring
De rol van hydrologie als sleutelfactor verraste ons, omdat we alleen een onderscheid hebben gemaakt tussen wel of geen afvoerloosheid en de stroomsnelheden hebben geschat. Van oudsher koppelen waterschappen hun ecologie-meetpunten aan meetpunten voor fysisch-chemische waterkwaliteit, maar onze studie heeft duidelijk gemaakt dat een koppeling met hydrologische meetpunten ook waardevol is om inzicht te krijgen in knelpunten voor waterflora. Zo kunnen op den duur wellicht drempelwaarden (in termen van duur van afvoerloosheid of stroomsnelheid) geformuleerd worden.
Ook alkaliniteit verdient een prominente plaats in het meetpakket van biologie-ondersteunende stoffen. Alkaliniteit is namelijk een sleutelfactor voor waterplanten (afbeelding 1). Bij veel waterschappen is alkaliniteit echter geen vaste parameter omdat hiervoor geen normen zijn geformuleerd. Tot slot is het wenselijk ook aspecten van de waterbodem, die op veel plaatsen nog opgeladen is met fosfaat [6], in de monitoring op te nemen.
Analyseren is leren!
Door onze eigen gegevens eens goed tegen het licht te houden hebben we scherper inzicht gekregen in onze mogelijkheden om de waterflora te versterken en ons meetnet te verbeteren. Wij raden andere waterschappen aan ook hun monitoringsgegevens niet alleen te uploaden, maar vooral ook zelf te analyseren. De informatie is te waardevol om ze alleen naar Brussel te versturen!
Dankwoord
Wij danken Annemarie Kramer-Hoenderboom, Marga Limbeek (beiden WRIJ) en Anita Buschgens (Ws V&V) voor hun stimulerende begeleiding van dit onderzoek.
Literatuur
- Puijenbroek, P. van (2014). De kwaliteit van het Nederlandse oppervlaktewater beoordeeld volgens de Kaderrichtlijn Water (KRW). Planbureau voor de Leefomgeving, publicatienummer 1355.
- Boedeltje, G. (2014). Waterplanten in relatie tot waterkwaliteit in de Achterhoek, Gelderse Vallei en op de Veluwe. Bureau Daslook, Waterschap Rijn en IJssel en Waterschap Vallei en Veluwe.
- Riis, T., K. Sand-Jensen, & O. Vestergaard (2000) . Plant communities in lowland Danish streams: species composition and environmental factors. Aquatic Botany, 66, 255-272.
- Hill, M.O. & P. Smilauer (2005). TWINSPAN for Windows, version 2.3. Centre for Ecology and Hydrology & University of South Bohemia, Huntingdon & Ceske Budejovice.
- Braak, C.J.F ter & P. Smilauer (1998). CANOCO Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination (version 4). Microcomputer Power Ithaca, New York.
- Lamers, L., S. Schep, J. Geurts & A. Smolders (2012). Erfenis fosfaatrijk verleden: helder water met woekerende waterplanten. H2O, 13, 29-31.
- Bloemendaal, F.H.J.L. & J.G.M. Roelofs (red.) (1988). Waterplanten en waterkwaliteit. Koninklijke Nederlandse Natuurhistorische Vereniging, Utrecht.
- Pot, R. (2014). QBWat, programma voor toetsing van opnamen aan de maatlatten van de KRW.
- Molen, D.T. van der, R. Pot, C.H.M. Evers & L.L.J. van Nieuwerburgh (red.) (2012). Referenties en maatlatten voor natuurlijke watertypen voor de Kaderrichtlijn Water 2015-2021. STOWA-rapport 2012-31.
- Best, E.P.H. (1976). Effect of nitrogen on the growth and nitrogenous compounds of Ceratophyllum demersum Aquatic Botany, 8, 197-206.
- Boedeltje, G., A.J.P. Smolders, L.P.M. Lamers & J.G.M. Roelofs (2005). Interactions between sediment propagule banks and sediment nutrient fluxes explain floating plant dominance in stagnant shallow waters. Archiv für Hydrobiologie, 162, 349-362.
- Lucassen, E.C.H.E.T. (2004). High groundwater nitrate concentrations inhibit eutrophication of sulphate-rich freshwater wetlands. Biogeochemistry, 67, 249-267.
- Evers, C.H.M., A.J.M. van den Broek, R. Buskens, A. van Leerdam, R.A.E. Knoben & F.C.J. van Herpen (2012). Omschrijving MEP en maatlatten voor sloten en kanalen voor de Kaderrichtlijn Water 2015-2021. STOWA-rapport 2012-34.
- Besozzi, D., D. ten Hulscher, M. Janssen, K. Maslowski, D.M. Saha, D. Schudoma, M. Wimmer & B. Zedler (2009). Afleiding van milieukwaliteitsnormen voor Rijnrelevante stoffen. Internationale Commissie ter Bescherming van de Rijn (ICBR), Koblenz.
- Cao, T., P. Xie, L. Ni, A. Wu, M. Zhang, S. Wu & A.J.P. Smolders (2007). The role of NH4+ toxicity in the decline of the submersed macrophyte Vallisneria natans in lakes of the Yangtze River basin, China. Marine and Freshwater Research, 58, 581-587.
- Boedeltje, G., A.J.P. Smolders & J.G.M. Roelofs (2005). Combined effects of water column nitrate enrichment, sediment type and irradiance on growth and foliar nutrient concentrations of Potamogeton alpinus. Freshwater Biology, 50, 1537-1547.
- Baggelaar, P., O. van Tongeren, R. Knoben & W. van Loon (2010). Rapporteren van betrouwbaarheid van de KRW-beoordelingen. H2O, 16, 21-25.















